
Кпд фотосинтеза в производственных посевах
Урожай создается в процессе фотосинтеза, когда в зеленых растениях образуется органическое вещество из диоксида углерода, воды и минеральных веществ. Энергия солнечного луча переходит в энергию растительной биомассы. Эффективность этого процесса и в конечном счете урожай зависят от функционирования посева как фотосинтезирующей системы.
В полевых условиях посев (ценоз) как совокупность растений на единице площади представляет собой сложную динамическую саморегулирующуюся фотосинтезирующую систему. Эта система включает в себя много компонентов, которые можно рассматривать как подсистемы; она динамическая, так как постоянно меняет свои параметры во времени; саморегулирующаяся, так как, несмотря на разнообразные воздействия, посев изменяет свои параметры определенным образом, поддерживая гомеостаз.
Такую систему характеризуют новые свойства по сравнению с отдельным растением. Так, для отдельного растения увеличение площади питания и связанное с этим улучшение освещенности приводят к повышению его семенной продуктивности, а для ценоза важна оптимальная густота растений. Условия для максимальной продуктивности отдельного растения и ценоза как системы не совпадают.
Исследованию фотосинтеза ценозов была посвящена Международная биологическая программа. Изучение фотосинтетической деятельности растений в посевах тесно связано с теорией получения высоких урожаев и возможностью управления формированием урожая. Методические основы ее изучения разрабатывали многие ученые. Большой вклад внес А. А. Ничипорович.
Управление формированием урожая весьма сложно, так как растения в ценозе, изменяясь в процессе вегетации, взаимодействуют с другими сложными системами — микроорганизмами почвы, возбудителями болезней, сорняками, вредителями. Многие факторы среды — температурный режим, осадки и др. — практически невозможно контролировать. Однако на основании анализа природно-климатических факторов можно подобрать сорта, адаптированные к конкретным условиям, разработать технологию их возделывания. Многие факторы поддаются регулированию. Можно изменить условия минерального питания, воздействовать на сорняки, вредителей и болезни. Управление процессом формирования урожая ведут на основе систематического контроля за развитием растений и направления хода фотосинтетической деятельности посевов в соответствии с заранее заданными параметрами.
Фотосинтетически активная радиация
Необходимое условие фотосинтеза — энергия солнечной радиации. Следует создавать такие посевы, в которых листья поглощали бы энергию солнца с возможно более высоким коэффициентом полезного действия для создания наибольшей биомассы и сосредоточения ее в хозяйственно ценной части урожая — семенах, клубнях, корнеплодах и т. п.
В процессе фотосинтеза принимает участие не вся солнечная энергия, а только ее видимая часть — фотосинтетически активная радиация (ФАР) с длиной волн от 380 до 720 нм (нанометр). Эти лучи поглощаются хлорофиллом и являются энергетической основой фотосинтеза. Энергия ФАР составляет около 50 % общей энергии солнечной радиации. Инфракрасная часть солнечного спектра, составляющая также около 50 % общей энергии солнца, не участвует в фотохимических реакциях фотосинтеза. Эти лучи поглощаются почвой, от которой нагреваются приземный слой воздуха и сами растения, при этом усиливаются транспирация и испарение влаги с поверхности почвы. Количество ФАР, падающее на единицу поверхности почвы в среднем по месяцам года и по декадам месяца, определено для различных географических зон и приведено в соответствующих справочниках.
Объективным показателем величины урожая (высокий, средний, низкий) может служить коэффициент использования ФАР. Хорошие урожаи соответствуют 2…3 % использования ФАР. При возделывании сортов интенсивного типа и оптимизации всех процессов формирования урожая возможна аккумуляция в урожае 3,5…5,0 % ФАР и более.
Поступление солнечной энергии за вегетационный период зависит от географической широты. Так, если сравнивать северные и южные районы России, то приход ФАР за возможный вегетационный период различается более чем в 2 раза. Кроме того, можно отметить, что на плодородных почвах Краснодарского края при достаточном количестве тепла и влаги значительно легче сформировать посев, который в среднем за вегетацию усваивает 3 % ФАР, чем в Тверской области на малоплодородных почвах и при недостатке тепла.
Показатели фотосинтетической деятельности посевов
Посев представляет собой оптическую систему, в которой листья поглощают ФАР. В начальный период развития растений ассимиляционная поверхность невелика и значительная часть ФАР проходит мимо листьев, не улавливается ими. С повышением площади листьев увеличивается и поглощение ими энергии солнца. Когда индекс листовой поверхности* составляет 4…5, т. е. площадь листьев в посеве 40…50 тыс. м2/га, поглощение ФАР листьями посева достигает максимального значения — 75…80 % видимой, 40 % общей радиации. При дальнейшем увеличении площади листьев поглощение ФАР не повышается.
В посевах, где ход формирования площади листьев оптимальный, поглощение ФАР может составить в среднем за вегетацию 50…60 % падающей радиации. Поглощенная растительным покровом ФАР — энергетическая основа для фотосинтеза. Однако в урожае аккумулируется только часть этой энергии. Коэффициент использования ФАР обычно определяют по отношению к падающей на растительный покров ФАР. Если в урожае биомассы в средней полосе России аккумулировано 2…3 % прихода на посев ФАР, то сухая масса всех органов растений составит 10… 15 т/га, а возможная урожайность — 4…6 т зерна с 1 га. В изреженных посевах коэффициент использования ФАР составляет всего 0,5…1,0%.
При рассмотрении посева как фотосинтезирующей системы урожай сухой биомассы, создаваемый за вегетационный период, или его прирост за определенный период зависит от величины средней площади листьев, продолжительности периода и чистой продуктивности фотосинтеза за этот период.
где У —урожайность сухой биомассы, т/га; ФП— фотосинтетический потенциал, тыс. м 2 • дни/га; ЧПФ —чистая продуктивность фотосинтеза, г/(м 2 • дни).
Фотосинтетический потенциал рассчитывают по формуле
где Sc — средняя за период площадь листьев, тыс. м 2 /га; Т — продолжительность периода, дни.
Основные показатели для ценоза, как и урожайность, определяют в расчете на единицу площади —1м 2 или 1 га. Так, площадь листьев измеряют в тыс. м 2 /га. Кроме того, пользуются таким показателем, как индекс листовой поверхности.
Динамика площади листьев в посеве подчиняется определенной закономерности. После появления всходов площадь листьев медленно повышается, затем темпы нарастания увеличиваются. К моменту прекращения образования боковых побегов и роста растений в высоту площадь листьев достигает максимальной за вегетацию величины, затем начинает постепенно снижаться в связи с пожелтением и отмиранием нижних листьев. К концу вегетации в посевах многих культур (зерновые, зерновые бобовые) зеленые листья на растениях отсутствуют.
Динамика площади листьев показывает, что на разных этапах вегетации посев как фотосинтезирующая система функционирует неодинаково (рис. 3). Первые 20…30 дней вегетации, когда средняя площадь листьев составляет 3…7 тыс. м2/га, большая часть ФАР не улавливается листьями, и поэтому коэффициент использования ФАР не может быть высоким. Далее площадь листьев начинает быстро нарастать, достигая максимума. Как правило, это происходит у мятликовых в фазе молочного состояния зерна, у зерновых бобовых в фазе полного налива семян в среднем ярусе, у многолетних трав в фазе цветения. Затем площадь листьев начинает быстро снижаться. В это время преобладают перераспределение и отток веществ из вегетативных органов в генеративные.

Рис. 3. Оптимальная площадь листьев в посевах однолетних культур с длительностью вегетационного периода 100 и 120 дней
На продолжительность этих периодов и их соотношение влияют различные факторы, в том числе агротехнические. С их помощью можно регулировать процесс нарастания площади листьев и продолжительность периодов. В засушливых условиях густоту растений, а следовательно, и площадь листьев намеренно снижают, так как при большой площади листьев усиливается транспирация, растения сильнее страдают от недостатка влаги, урожайность уменьшается.
Слишком большое разрастание площади листьев при достаточном водоснабжении также приводит к нежелательным результатам. Биомасса в этом случае растет довольно высокими темпами за счет вегетативных органов, однако условия формирования плодов и семян ухудшаются. К подобным результатам может привести и чрезмерное загущение растений. Для кормовых культур, у которых листья представляют хозяйственно ценную часть урожая (например, травы), площадь листьев может достигать 60…80 тыс. м 2 /га.
Таким образом, получение высоких урожаев связано с оптимальным ходом (графиком) нарастания площади листьев. Такие графики могут быть определены для каждой культуры и сорта в конкретных условиях их выращивания.
Формирование урожая зависит не только от величины площади листьев, но и от времени ее функционирования. Фотосинтетический потенциал (ФП) объединяет эти показатели. ФП может быть определен за любой период времени, например за декадные, межфазные периоды или в целом за вегетационный период. ФП за какой-либо период представляет сумму величин площади листьев за каждые сутки периода. Например, если площадь листьев в начале периода составляла 20 тыс. м 2 /га, а через 10 дней — 28 тыс. м 2 /га, то ФП этого 10-дневного периода составляет (20 + 28): 2 • 10 = = 240 тыс. м 2 • дни/га. ФП хорошо развитых посевов зерновых культур с вегетационным периодом 100…ПО дней составляет за вегетацию 2,0…2,5 млн м 2 • дни/га. В южных районах для культур с длительным вегетационным периодом и при хорошем водоснабжении ФП может составить до 4 млн м 2 • дни/га.
Чистая продуктивность фотосинтеза (ЧПФ) характеризует интенсивность фотосинтеза посева и представляет собой количество сухой массы растений в граммах, которое синтезирует 1 м 2 листовой поверхности за сутки. В среднем за вегетацию у таких культур, как пшеница, ячмень, ЧПФ составляет 5…7 г/(м 2 • дни). У кукурузы ЧПФ обычно выше. ЧПФ, так же как и ФП, определяют за какой-либо период или в среднем за вегетацию:
где В2 и B1 — сухая масса растений с единицы площади в конце и в начале периода.
ЧПФ варьирует в течение вегетации. В первый месяц вегетации ЧПФ выше, чем в последующий, так как в начале вегетации растения не затеняют друг друга, все листья хорошо освещены. В дальнейшем с увеличением площади листьев ЧПФ начинает уменьшаться в связи с затенением нижних листьев.
В начале вегетации нарастание биомассы идет медленно, затем темпы приростов увеличиваются. В конце вегетации, когда площадь листьев небольшая, суточные приросты биомассы также невелики. В это время идет перераспределение накопленных ассимилятов из листьев, стеблей и корней в генеративные органы.
Прирост биомассы за любой промежуток времени, в том числе и за вегетацию, равен произведению ФП и ЧПФ. Если в среднем за 100 дней вегетации ЧПФ равнялась 6 г/(м 2 • дни), а ФП — 2 млн м 2 • дни/га, то количество сухой биомассы составит 12 т/га.
Посев как фотосинтезирующая система наиболее производительно функционирует в период, когда площадь листьев близка к оптимальной — 30…50 тыс. м 2 /га. Если ЧПФ в это время равна 5…7 г/(м 2 •дни), то при площади листьев 40 тыс. м 2 /га суточный прирост сухой биомассы составит 200…280 кг/га. Если период с такой средней площадью листьев продолжается 30 дней, то за это время прирост сухой биомассы составит 6…8 т/га. Прирост биомассы за период, когда посев функционирует в оптимальном режиме, составляет более 70 % максимального за вегетацию, хотя продолжительность этого периода — всего 30 % общей вегетации культуры. Коэффициент использования ФАР в это время в 2…3 раза больше, чем в первый месяц после появления всходов, а также в конце вегетации.
При созревании в корнях и стеблях сосредотачивается 50…60 % сухой массы растений, в основном представленной клетчаткой. В нашем примере из общей биомассы 12 т/га на урожай зерна приходится 5…6 т.
Различия в урожаях в конечном итоге определяются тем, достигал ли и как скоро достигал посев в своем развитии оптимального для данных условий состояния, а также как долго он функционировал в этом состоянии.
Таким образом, высокие урожаи обеспечиваются определенным ходом фотосинтетической деятельности растений в посевах. Оптимальный ход нарастания площади листьев и биомассы может быть определен для каждой культуры и сорта в конкретных условиях выращивания.
Факторы, лимитирующие фотосинтез
Почему растения в посеве не достигают оптимального уровня показателей фотосинтетической деятельности или существенно отклоняются от них в отдельные периоды вегетации? Какие элементы фотосинтетической деятельности подвержены наибольшему влиянию различных факторов?
Как следует из формулы У = ФП • ЧПФ, урожай биомассы (а также и хозяйственно ценной части урожая) прямо зависит от этих показателей. При улучшении условий жизни растений (оптимизация режима питания и влагообеспеченности) обычно усиливаются ростовые процессы, увеличивается площадь листьев. В этом случае листья сильнее затеняют друг друга, поэтому ЧПФ снижается.
Большинство культурных растений относятся к типу С3, у которых фотосинтез усиливается с ростом концентрации СO2 при насыщающей интенсивности освещения.
К растениям типа С4 относятся хлеба второй группы — кукуруза, сорго, просо, рис и сахарный тростник. У них не наблюдается светового насыщения и усиленного фотодыхания, а компенсационная точка по СO2 необычайно низка. ЧПФ у С4-растений выше, чем у С3-растений, особенно при повышенной площади листьев. Сорта детерминантного типа роста (ультраскороспелые сорта сои) обычно характеризуются более высокой ЧПФ. Имеют значение также форма, направление и анатомическое строение листьев. У сортов люпина узколистного ЧПФ обычно выше, чем у сортов других видов люпина.
Считается, что у зерновых культур фотосинтез посева осуществляется лучше, если верхние листья направлены под острым углом к стеблю. В процессе селекционного совершенствования сахарной свеклы распластанная по поверхности почвы розетка листьев постепенно превращалась в воронкообразную, что определило способность современных сортов создавать высокопродуктивные посевы.
Большое значение имеет аттрагирующая способность генеративных и запасающих органов растений, благодаря которой пластические вещества из листьев активно перемещаются в них.
Для скороспелых форм ФП должен составлять 1,5…2,0 млн м 2 • дни/га. для среднеспелых — 2,5…3,0 и для позднеспелых — 3…5 млн м 2 • дни/га.
В пределах одного типа скороспелости увеличение ФП может происходить путем ускорения темпов роста площади листьев и увеличения ее максимального значения. Густота посевов —один из факторов, сильно влияющих на эти показатели. Однако чрезмерное разрастание листьев при усилении их затенения может стать отрицательным фактором для формирования репродуктивных органов.
При оптимальной влагообеспеченности фактором, ограничивающим ростовые процессы, становится уровень минерального питания. Для получения высоких урожаев необходима высокая обеспеченность элементами минерального питания, в первую очередь азотом. Так, растения пшеницы при урожайности зерна 5 т/га выносят из почвы 170…200 кг азота с 1 га. Однако высокие нормы азота при достаточном количестве влаги усиливают вегетативный рост растений и разрастание листовой поверхности сверх оптимальных размеров, растения полегают, урожай снижается.
Основное средство повышения эффективности использования ценозом влаги и удобрений на фотосинтез и повышение урожайности — генетическое преобразование растений, создание сортов, у которых рост вегетативных органов ограничен, а генеративных — значительно выше, чем у старых сортов. Поэтому одно из направлений современной селекции — создание сортов, реагирующих на улучшение условий выращивания увеличением хозяйственно ценной части урожая.
В онтогенезе растений от всходов до начала созревания выделяют 4 биологически обоснованных периода:
- всходы — начало цветения;
- цветение и образование плодов;
- рост плодов;
- налив семян.
Основные выходные показатели первого периода — площадь листьев и ФП; второго — максимальная за вегетацию площадь листьев, ФП этого периода и количество образовавшихся плодов в расчете на единицу площади (на 1 м 2 ). Число образовавшихся плодов прямо коррелирует с ФП первого периода и особенно тесно — с ФП второго.
В третий период (рост плодов) площадь листьев постепенно начинает уменьшаться, но в среднем сохраняется на высоком уровне; биомасса продолжает интенсивно нарастать за счет роста плодов, которые к концу периода достигают максимальной величины. От числа сохранившихся к концу третьего периода плодов и семян зависят показатели фотосинтеза и поступление в растения азота в четвертый период, когда происходит налив семян. Посев как фотосинтезирующая система наиболее активно функционирует в течение второго и третьего периодов. За это время, равное по продолжительности первому периоду, накапливается 60…70 % биомассы и такое же количество азота от максимальных за вегетацию величин.
Таким образом, эффективность работы каждого последующего периода зависит не только от агрометеорологических условий этого периода, но и от результатов функционирования посева в предыдущий период. Конечные показатели развития посева — урожай семян и его качество — зависят от последовательных состояний посева в процессе роста и развития растений.
При этом каждый квадратный метр листовой поверхности будет работать с несколько сниженной интенсивностью фотосинтеза, но зато с повышенным КПД, а идеальный по структуре посев с достаточно большой площадью листьев сможет проводить наибольшую суммарную фотосинтетическую работу, т. е. лучше всего использовать приходящуюся на него энергию солнечной радиации.
Чтобы подчеркнуть значение посевов и ценозов как целостной фотосинтезирующей системы, при помощи которой можно использовать приходящую энергию солнечной радиации с высоким КПД, приведем такой пример: представим себе, что растения посева образуют один сплошной слой горизонтальных, примыкающих друг к другу листьев (монослой). Площадь их в сумме будет составлять 10 000 м 2 /га.
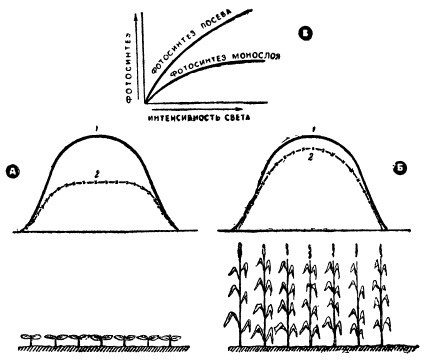
Стремясь к повышению коэффициента использования энергии солнечной радиации в процессе фотосинтеза посевов, мы должны стремиться к решению двух основных задач:
- Всемерно улучшать интенсивность фотосинтеза отдельных листьев растений. (На графиках такой процесс выразится в повышении крутизны подъема световых кривых и уровня плато светового их насыщения).
- Создавать такие посевы, которые использовали бы с высоким КПД свет как низкой, так и высокой интенсивности, то есть обладали бы в целом световыми кривыми фотосинтеза, наиболее круто поднимающимися и имеющими высокий уровень плато.
Полноценный сомкнувшийся посев с хорошими световыми кривыми может использовать на фотосинтез поглощаемую за день энергию радиации с КПД, доходящим до 15%, а приходящую (имеется в виду поглощение ее в количестве 85%) — около 12—13%. Это хороший КПД использования энергии солнечной радиации.
Здесь описана возможная эффективность фотосинтетической работы посева в наилучшем состоянии, т. е. в таком, когда он обладает большой площадью листьев и хорошей структурой. На самом деле в таком состоянии даже самые хорошие посевы бывают только часть Бремени из общего периода произрастания. Значительная же часть времени уходит на появление всходов, на первые этапы их развития. В это время травостои еще не смыкаются и значительная часть энергии солнечной радиации падает на почву, безвозвратно теряясь для фотосинтеза.
После периода оптимального состояния начинается период созревания растений. Листья теряют фотосинтетическую активность. Формирование хозяйственной части урожая идет не столько за счет фотосинтеза, сколько за счет перемещения ранее накопленных пластических веществ из листьев, стеблей в репродуктивные и запасающие органы: в зерно, клубни, корнеплоды и т. д.
В конце концов, в сумме посев за время своего развития использует энергию солнечной радиации на фотосинтез с КПД более низкими, чем указано выше. При этом в разных случаях и в разных посевах показатели суммарного итогового КПД могут изменяться очень сильно. Со всем этим приходится считаться. Но даже при всех трудностях есть немало способов направлять ход формирования посевов по наилучшим путям и добиваться высоких КПД использования ими энергии на фотосинтез.
Для успешного достижения желаемых результатов надо иметь полное представление о взаимосвязях и предпосылках, которые лежат в основе формирования фотосинтезирующих систем, способных осуществлять фотосинтез с наиболее высокими КПД.
Об этих предпосылках и принципах можно наиболее доступно рассказать на примере действия фотосинтезирующих систем в виде суспензий одноклеточных водорослей.
План лекции:
1. Урожай как результат фотосинтетической деятельности растений в посевах;
2. Фотосинтез как фактор урожайности;
3. Фотосинтетически активная радиация (ФАР);
4. Фактические и теоретически возможные коэффициенты использования солнечной энергии;
5. Получение посевов с оптимальным ходом роста площади листьев;
6. Ресурсы ФАР и потенциальный урожай;
7. Тепло как часть солнечной радиации.
1. Основная литература
1.1 Баранов В. Д., Тараканов И. Г. Программирование урожаев сельскохозяйственных культур.-М.: Изд-во УДН, 1990. – 71 с.
1.2 Гаврилов А. М. и др. Программирование урожаев сельскохозяйственных культур. - Волгоград: ВСХИ, 1984. - 194 с.
1.3 Каюмов М. К., Программирование продуктивности полевых культур. Справочник. М: Росагропромиздат, 1989. - 368 с.
1.4 Листопад Г. Е. Программирование урожаев. - Волгоград. 1975. - 368 с.
2. Дополнительная литература
2.2.Собого А. А. и др. Программирование урожаев - в основу прогрессивных технологий. - К.: Урожай, 1984. - 150 с.
2.3.Томин Х. Г. Солнечная радиация и формирование урожая. - Л.: Гидрометеоиздат, 1977. - 197 с.
1. Урожай как результат фотосинтетической деятельности растений в посевах
Физиология растений дала ряд критериев в определении высокого урожая. Растениеводство является системой мероприятий, направленных на наилучшее использование процесса фотосинтеза зеленых растений.
Повышение валовых сборов продукции возделываемых растений возможно осуществить двумя путями. Первый путь - это расширение посевных площадей. В этом случае возможности ограничены, так как проведенные подсчеты показывают, что в результате мелиорации и освоения ряда территорий площади под сельскохозяйственными растениями могут быть увеличены в 2-3 раза. Увеличение же посевов на освоенных площадях одной культуры связано с сокращением посева другой.
Второй путь - повышение урожайности благодаря увеличению Продуктивности фотосинтеза растений в посевах и, в частности, повышение чистой продуктивности фотосинтеза. Этот путь увеличения сборов продукции через фотосинтез таит в себе большие возможности, так как 90-95% биомассы растений составляют органические вещества, образуемые в процессе фотосинтеза. В то же время выяснено, что конечным решающим фактором, определяющим максимально возможную урожайность, может быть приход солнечной радиации.
Конкретной задачей на ближайшие годы в этой области является нахождение условий для повышения фотосинтетической активности растений в посевах, доведя использование ими солнечной энергии на фотосинтез вместо 0, 5-1% падающей на поле до 3-5%, что соответственно повысит и накопление органической массы.
Главная особенность процесса фотосинтеза состоит в том, что, используя воду как источник водорода, зеленые растения переносят с помощью энергии солнечного света (ФАР 380-710 нм) водород на восстановление СО2, и в результате этого образуются свободный кислород и органические вещества, в частности углеводы (условно СН2О):
Однако в процессе фотосинтеза образуются не только углеводы. В общий метаболизм вовлекаются также азот, фосфор, сера, магний, калий, железо, медь, молибден и другие элементы минерального питания. Многие из них участвуют в образовании неуглеводных продуктов фотосинтеза (аминокислот, белков, липидов, хлорофилла), в биосинтезе структур и агентов самого фотосинтетического аппарата (Ничипорович, 1977).
Фотосинтез является и первоисточником той биологически активной энергии, которая необходима для усвоения элементов минерального питания. Решающая роль фотосинтеза в формировании урожаев определяется усвоением углерода и энергии, а также, прямо или косвенно, и элементов минерального питания растений.
Биологический предел продуктивности листа растений или посева может быть достигнут тогда, когда фотосинтез будет осуществляться с максимально возможным коэффициентом использования приходящей энергии ФАР.
Таким образом, показатели КПД ФАР в формировании биологических урожаев являются одними из важных критериев для оценки достигаемых уровней активности и в разработке принципов и приемов ее увеличения.
2. Фотосинтез как фактор урожайности.
Проблема фотосинтеза и использования солнечной радиации - одна из важнейших в международной биологической программе. В настоящее время существуют два направления в изучении процесса фотосинтеза.
С позиции физики, химии и физической химии познание процесса фотосинтеза может привести к синтезу ценных органических продуктов путем использования даровых источников сырья - углекислого газа, карбонатов, воды и энергии солнца.
С физиологической и экологической точек зрения исследователи имеют целью всестороннее и наиболее полное изучение фотосинтеза как физиологического процесса, чтобы научно обосновать приемы возделывания растений, обеспечивающие наилучшие условия питания, и получение максимальных урожаев продукции высокого качества.
Л. А. Иванов выразил соотношение между фотосинтезом и урожаем, следующим уравнением:
М + m = FРТ-АР1Т1
где М - прирост массы сухого вещества растений за учитываемый период; m - масса отмерших за время вегетации частей растений;
F - интенсивность фотосинтеза;
А - интенсивность дыханий;
Р - фотосинтезирующая площадь;
Т - продолжительность процесса фотосинтеза;
Р1 - рабочая поверхность дыхания;
Т1 - продолжительность процесса дыхания.
Показатель FРТ в этом уравнении, представляющий собой произведение интенсивности фотосинтеза, величины фотосинтетической (листовой) поверхности и продолжительности процесса фотосинтеза, назван продуктивностью фотосинтеза и определяет урожай сухой массы.
Большой размер урожая и высокое его качество могут быть получены только в том случае, если суммарный фотосинтез будет перекрывать затраты пластических материалов на интенсивные ростовые процессы и дыхание. В этом случае возникнут некоторые излишки ассимилянтов, являющихся источником повышения сахаристости, крахмалистости и т. д. Для выражения этой зависимости был введен показатель - коэффициент эффективности фотосинтеза:

В процессе фотосинтеза в растении образуются и накапливаются органические вещества. Количество их зависит от интенсивности фотосинтеза и дыхания растений, от соотношения между этими процессами, от положения так называемой компенсационной точки. Это такое состояние, когда в растении фотосинтез и дыхание полностью уравновешиваются, оно означает, что сколько вещества создано, столько и израсходовано в процессе дыхания, т. е. нет ни увеличения, ни уменьшения массы, окружающая атмосфера не обогащается ни кислородом, ни углекислым газом. При таких условиях органические вещества не накапливаются.
Накопление органического вещества растением за определенный период времени или за всю его жизнь следует рассматривать как разницу между количеством созданного на свету органического вещества и израсходованного на дыхание. Кроме того, во время вегетации в растении могут происходить превращения продуктов фотосинтеза с изменением их массы. Например, масса откладываемой в запас клетчатки на 10% меньше, чем масса глюкозы, из которой она образуется. К этому следует добавить, что отмирание корневых волосков и мелких корешков, сбрасывание части цветков — потери, происходящие в период вегетации. Поэтому общая продуктивность растения будет определяться не только образованием органических веществ в процессе фотосинтеза, но и величиной всех указанных потерь.
Доказано, что энергия, используемая для образования продуктов фотосинтеза при благоприятных условиях (высокой концентрации СО2 в окружающем воздухе, хорошем водоснабжении и т. д.), составляет всего 2-5%. Обычно при высокой интенсивности радиации количество энергии, затраченной на фотосинтез, равно 1%, или 16 кДж/м 2 в 1 ч.
На основании ряда исследований, в частности работ английских ученых Броуна, Эскомба и Вильсона, можно утверждать, что при нагревании листа растение может отдавать в виде теплоизлучения 84 кДж/м 2 в 1 ч на каждый градус температурного градиента между листом растения и окружающим воздухом. Остальная энергия в количестве 1500 кДж/м 2 в 1 ч (99% энергии, или 1584 кДж без энергии теплоизлучения), превращаясь в тепло, способствует нагреванию и испарению воды из листьев — транспирации, требующей большого расхода энергии. На испарение 1 г воды при 20°С затрачивается 2,45 кДж (скрытая теплота испарения воды). Растение, поглощая 1500 кДж/м 2 в 1 ч, может испарить 612 г воды, что соответствует транспирационному коэффициенту ряда растений.
Подсчеты коэффициента использования солнечной энергии показали, что максимальный прирост сухого вещества на одно растение кукурузы в период выбрасывания метелки за сутки составляет 20 г. Принимая это сухое вещество за крахмал, что почти соответствует действительности, и зная, что теплота сгорания 1 г крахмала составляет 16,6 кДж, можно рассчитать суточный прирост энергии на одно растение кукурузы, который будет составлять 351 кДж. На 1 га при площади питания 70×70 см размещается 30-40 тыс. растений кукурузы. Таким образом, за сутки накопится 10534×10 3 —14045×10 3 кДж энергии. За 6 ч на поверхность 1 га посевов кукурузы в летний период поступает 209 млн кДж, из них растениями кукурузы используется лишь 5-6% энергии солнечной радиации. Расчеты показали, что на создание урожая яровая пшеница использует 3,26%, картофель — 3,02, сахарная свекла — 2,12% лучистой энергии.
Количество ФАР, поглощенной посевом, растительным покровом, определяют по формуле, Дж/см 2 в 1 мин: $$\large \Pi = Q - R - T_n + R_n$$
где П — поглощенная посевом радиация; Q — суммарная радиация, падающая на посев; R — радиация, oтраженная от посева и вышедшая за пределы его верхней границы; Тn — радиация, проникшая к почве; Rn — радиация, отраженная от почвы под растительностью.
Коэффициент поглощения энергии ФАР (Qп) посевом определяют делением обеих частей формулы на Q: $$\large Q_\pi = \frac<\Pi> = 1 -\frac - \frac + \frac, $$
где \(\frac\) — альбедо посева, показывающее, какая доля падающей радиации у отражается посевом;\(\frac\) — коэффициент пропускания, показывающий, какая доля падающей радиации (Q) достигает почвы под растительностью; \(\frac\) — альбедо почвы под растительностью (К. Г. Тооминг, Б. Н. Гуляев).
Эффективность фотосинтеза можно характеризовать коэффициентом полезного действия (КПД), который определяют по формуле: $$\large E \% (КПД) = \frac,$$
где А — количество энергии, поступившей за период вегетации па 1 га посева, или энергии, которая была поглощена посевом, кДж; В — количество энергии, накопившейся в органической массе урожая (биологического или хозяйственного), кДж.
В среднем КПД фотосинтеза сельскохозяйственных растений составляет 0,5-1%, а теоретически возможный — 4-6%. Максимальный показатель использования энергии ФАР посевами и насаждениями обусловливается способностью поглощать не менее 60% энергии света, поступающего к ним на протяжении вегетации. Из этого количества на фотосинтез идет 10%, на дыхание расходуется около 20% энергии, усваивающейся в процессе фотосинтеза, или $$\large E \% (КПД) = 60 \cdot \frac \cdot \frac = 48 \%.$$
В 1 кг сухой массы урожая накапливается до 17 тыс. кДж энергии.
Поступающая солнечная энергия в разных зонах неодинаково взаимодействует с режимом температуры и влаги, вследствие чего значительно изменяется эффективность солнечной радиации как фактора фотосинтеза и продуктивности. Кроме того, следует отметить, что она является не только движущей силой фотосинтеза, но и основным фактором транспирации. Свет влияет и па такие физиологические процессы, как рост, органогенез, передвижение ассимилятов. Один из основных путей повышения продуктивности фотосинтеза — увеличение до определенных размеров площади ассимилирующих органов — листьев и усиление их деятельности, что обусловливает количество лучистой энергии, поглощаемой хлоропластами. Доказано, что величина урожая в значительной мере зависит от оптималык структуры посевов. Структурой посевов называется создаваемая архитектоника (построение) сообщества растений, которое характеризуется определенными морфологическими признаками и физиологическими функциями, а оптимальная структура — это такой посев, который имеет высокий КПД фотосинтеза и обеспечивает максимальный биологический и хозяйственный урожай.
| Вариант | Дни после всходов | ||||||||
| 27 | 36 | 46 | 59 | 68 | 87 | ||||
| Площадь листьев, тыс. м 2 | |||||||||
| Без подкормки | 6,6 | 11,4 | 17,2 | 19.8 | 19,8 | 19,3 | |||
| Подкормлено NPK | 7,4 | 11,8 | 20,3 | 21.9 | 21,3 | 19,6 | |||
| Поглощено ФАР, тыс. кДж/мин | |||||||||
| Без подкормки | 11,0 | 84,9 | 109,3 | 130,8 | 141,7 | 137,5 | |||
| Подкормлено NPK | 68,1 | 91,1 | 139,6 | 157,2 | 163,9 | 149,2 | |||
Установлено, что при подкормке растений увеличиваются размеры листовой поверхности, а также изменяются физиологические особенности фотосинтетического аппарата— способность поглощать и усваивать лучистую энергию. Так, более развитая листовая поверхность подкормленных растений кукурузы в период вегетации поглощала и большее количество ФАР (табл. 10). Кроме того, листья кукурузы разных ярусов неодинаково поглощают лучистую энергию. Распределение ФАР внутри посева непропорционально площади листьев по отдельным ярусам. Например, верхний слой листьев составляет лишь 23,7% всей листовой поверхности, а поглощает 47% энергии, в то время как средний слой, составляющий 60,4% этой поверхности, усваивает только 36,6% поглощенной посевом ФАР. В результате подкормки растений NPK уменьшилось количество энергии, поглощаемой нижним ярусом листьев, с 26,4 до 8,6% и увеличился процент поглощения ФАР средним слоем — с 36,6 до 49,7%. Таким образом, условия минерального питания в значительной мере влияют на структуру посевов и поглощение ФАР листьями разных ярусов (слоев травостоя).
Повышение фотосинтетической продуктивности растений при бесперебойном снабжении их водой и питательными веществами обеспечивается наиболее целесообразным размещением растений на площади, т. е. так, чтобы они в достаточной степени освещались солнечными лучами. Поэтому в сельскохозяйственной практике используют различные способы посева. Для лучшего освещения растений, а также для более эффективного использования лучистой энергии рядки располагают с востока на запад или с северо-востока на юго-запад.
Таким образом, в течение вегетации необходимо контролировать и регулировать интенсивность фотосинтеза, энергетический баланс посева, КПД фотосинтеза за короткие промежутки времени, прирост общей сухой массы. В связи с этим поставлена задача создавать и внедрять в производство сорта, способные развивать большую листовую поверхность, имеющие высокую продуктивность фотосинтеза и дающие большой биологический и хозяйственный урожай.
Предположим, что интенсивность фотосинтеза будет не 10-15 мг, а 30-50 мг СО2 на 1 дм 2 в 1 ч, тогда при средней площади листьев 30-40 тыс. м 2 на 1 га у таких растений, как сахарная свекла, картофель, урожай корне- и клубнеплодов, будет составлять 6-8 т с 1 га (в пересчете на сухую массу).
Абсолютная величина продуктивности растения равна в среднем 40-50 мг органического вещества в сутки на 50 см 2 листовой поверхности при условии, что дыхание составляет не более 5-10% фотосинтеза. Фотосинтез в естественных условиях — процесс очень изменчивый, и по основным его показателям — интенсивности (мг СО2 на 1 дм 2 в 1 ч) и чистой продуктивности (г/м 2 в сутки) — наблюдается большая вариабельность. Чистую продуктивность фотосинтеза (Фч.пр), которая характеризует интенсивность образования и накопления органической массы урожая, определяют по формуле, г/м 2 : $$\large Ф_ = \frac<(Л_1 + Л_2) \frac<1>n> ,$$
Если, кроме общей чистой продуктивности фотосинтеза, надо определить интенсивность работы листьев и ее направленность на формирование хозяйственно ценных органов, например корнеплодов, клубней или зерна, в уравнение вместо В2 — B1 подставляют величину Х2 — X1, т. е. прирост сухой массы хозяйственно ценной части урожая.
Фотосинтез посевов (Фп) можно оценить, используя значение суточных приростов сухой фитомассы по формуле, предложенной Б. И. Гулиевым и А. С. Оканенко: $$\large Ф_n = \frac,$$
Например, для посева кукурузы получены такие значения (мг С02 на 1 дм 2 в 1 ч): в Киеве — 54, в Волгограде — 84, в Москве — 18.
Потенциальную фотосинтетическую мощность посева может характеризовать фотосинтетический потенциал растений — сумма ежедневных показателей площади листьев посева за весь вегетационный период или часть его, выраженная в м 2 дней/га.
Различают урожай биологический и хозяйственный.
Биологический урожай — это сумма суточных приростов за весь вегетационный период. Его можно выразить формулой $$\large У_ = \Sigma C_,n.$$
где С — суточные приросты массы, кг/га в сутки. $$\large C = \fracЛ>,$$
где Ф — количество ассимилированного СО2; Кэф — коэффициент фотосинтеза $$\large \left ( \frac>\; CO_2> \right);$$
Л — листовая поверхность; 1000 — граммы.
Хозяйственный урожай составляет определенную долю биологического. Не все части растения равноценны и одинаково используются. Например, у хлебных злаков наиболее ценным является зерно, у сахарной свеклы — корень и т. д. Таким образом, у различных культур коэффициент хозяйственного использования (Кхоз) разный. Поэтому хозяйственный урожай равен: $$\large У_ = У_К_.$$
Таким образом, дальнейшее повышение урожайности сельскохозяйственных культур должно осуществляться путем увеличения продуктивной части растения. Знание закономерностей, определяющих те или иные изменения интенсивности и продуктивности фотосинтеза, умение управлять этими изменениями — одна из важных основ получения больших биологических и хозяйственных урожаев высокого качества.
Продуктивность фотосинтеза сельскохозяйственных культур целесообразно оценивать величиной выхода полезной энергии с одного гектара.
Полезная энергия — это материализованная солнечная энергия в урожае за вычетом технологической энергии, затраченной на его производство (обработка почвы, удобрение, сорт, приемы агротехники).
При интенсивной технологии возделывания сельскохозяйственных культур, предусматривающей высокий КПД фотосинтеза и высокую урожайность, минимальные затраты технологической энергии, количественное соотношение содержащейся в урожае и технологической энергии может служить важным показателем энергетической эффективности культуры, т. е. энергетических затрат на производство единицы сельскохозяйственной продукции.
Фотосинтез растений изучается очень активно. В нашей стране ведутся широкие исследования по физико-химическим и прикладным проблемам этого процесса.
Познание природы фотосинтеза растений — одна из наиболее важных, фундаментальных проблем биологии, тесно связанных с удовлетворением нужд человечества в пище и энергии.
Значительные результаты достигнуты в изучении молекулярных механизмов преобразования энергии света при фотосинтезе. Установлено, что этот процесс начинается с поглощения энергии света (фотонов) пигментами антенны и миграции энергии возбуждения к реакционным центрам, где за пикосекунды происходит преобразование ее в химическую энергию; изучаются улътрамолекулярная структура реакционного центра и функции его компонентов; решается задача построения искусственных реакционных центров; большое внимание уделяется изучению механизма переноса электронов в фотосинтезирующих системах.
Ведутся исследования по направленному синтезу продуктов фотосинтетического метаболизма (углеводов, белков, жиров). Большое внимание уделяется применению принципов фотосинтеза для построения искусственных систем, моделей, использующих энергию света. Важное значение придается созданию биотехнических систем, потребляющих солнечную энергию для образования водорода и восстановления молекулярного азота, а также созданию фотобиотехнических систем, использующих солнечную энергию с предельно достижимым КПД (до 30%). Исследуются молекулярные фотокаталитические системы как преобразователи солнечной энергии при высоких концентрациях СО2 и высоких интенсивностях света.
По комплексной программе ведутся исследования бактериального фотосинтеза в направлении молекулярной организации фотосинтетических систем, поглощения, переноса и конверсии энергии света.
Выполнение всех этих и ряда других фундаментальных исследований внесет значительный вклад в теорию и практику фотосинтеза.
Читайте также: